
Executive summary
Fungal infections represent a significant and growing threat to global health, affecting millions of individuals annually. These infections range from superficial conditions, such as athlete's foot and oral thrush, to severe, life-threatening systemic diseases like invasive candidiasis, aspergillosis, and cryptococcosis.
Several factors are fueling an increase in fungal disease incidence and complications with treatment. Climate change and healthcare-associated infections are changing emergence patterns, increasing the opportunity for fungal infections. The cost of new drug development is also slowing down the process of finding new clinical treatments for these infections. While research would normally increase at this stage, we’re seeing publications and clinical trials level off and decline.
Fortunately, many innovative approaches are available to improve fungal disease treatment. AI and genetic editing are important new treatment options. Drug repurposing and combination therapies are leveraging existing medications for better effectiveness. Biomaterials and improved delivery methods are also providing potential new approaches. With renewed interest from researchers and the pharmaceutical industry, we can implement breakthroughs in treating fungal infections and stave off worsening conditions in the incidence of these infections for patients worldwide.
Introduction: The growing problem of fungal diseases
Fungal diseases constitute a substantial yet frequently overlooked global health threat, affecting approximately 6.5 million individuals annually, leading to over 3.8 million deaths. Unlike most infectious diseases, fungal infections primarily result from environmental exposure, commensal overgrowth, or reactivation of latent infection. In 2022, the World Health Organization (WHO) released its first-ever classification of fungal pathogens into critical, high, and medium priority categories, which underscores the growing recognition of this threat and aims to focus global response efforts on combating it.
Multiple factors are driving significant epidemiological shifts in fungal diseases. Modern medical advances, for example, have created paradoxical effects — prophylactic antifungal use has reduced candidemia incidence in certain populations while promoting resistance development. The expanding use of immunosuppressive medications, chemotherapeutic regimens, and antibiotics has created new and growing at-risk populations. Improved therapeutics for previously fatal conditions have extended patient survival, providing more opportunities for fungal infections to develop. In addition, the increased deployment of invasive medical devices and procedures offers new pathways for commensal fungi to access tissues and bloodstream.
Climate change is also playing a critical role in spreading fungal diseases (see Figure 1). As temperatures rise, previously inhospitable regions for various fungi are becoming more suitable, thereby extending their range to previously unexposed populations. A compelling example of this climate-driven range expansion is Coccidioides, the fungus responsible for Valley Fever. Historically confined to hot, arid environments of the southwestern United States, Mexico, and parts of Central and South America, this pathogen has demonstrated concerning northward migration. In 2015, researchers documented the presence of Coccidioides in Washington state, a region previously considered too cool for this fungus to establish itself.
Higher temperatures also drive evolutionary adaptations in fungi, enabling them to develop enhanced thermotolerance and overcome the thermal barrier that has historically protected humans from fungal infections. Candida auris exemplifies this concerning trend — in the U.S., its prevalence increased by over 200% between 2019 and 2021.
Climate change-driven natural disasters, such as floods and wildfires, significantly increase fungal infection risks. Floods create ideal environments for fungal growth because they generate widespread moisture that is essential for spore germination. Fires aerosolize fungal spores that can travel significant distances, exposing populations far from the original burn sites. This mechanism has triggered multiple Coccidioidomycosis outbreaks among firefighters and distant communities.
Not only does climate change directly expose humans to more fungal pathogens, but it also drives antifungal resistance via overuse of these treatments on agricultural fields. Rising temperatures enhance fungal virulence, compelling greater agricultural fungicide application to protect crops. This widespread deployment of agricultural azoles, particularly in warming regions, has emerged as a critical driver of resistance development.
Research demonstrates that the combination of heat stress and azole exposure creates ideal conditions for accelerated resistance evolution in fungal pathogens such as Aspergillus fumigatus, Candida species, and Cryptococcus species. The recent identification of Rhodosporidiobolus fluvialis, capable of temperature-induced mutagenesis, provides further evidence that rising global temperatures may accelerate the evolution of resistance and hypervirulence in fungi, presenting substantial challenges to clinical management in our warming climate.

Further complicating the issue of fungal infections is the nature of antifungal drugs, which can cause off-target effects and harm the host (i.e., the patient). Fungi cells share many similarities with human cells, making it more challenging to develop antifungal drugs that selectively target the fungal pathogen.
This has limited the number of antifungal drug classes available, with only a handful of agents such as polyenes, azoles, echinocandins, and allylamines forming the backbone of antifungal therapy. Each class targets specific fungal cellular components, such as the cell membrane (e.g., ergosterol) or cell wall (e.g., β-glucan), but their use is often constrained by issues such as toxicity, drug-drug interactions, and the emergence of resistance.
Finally, these challenges are compounded by the fact that developing new antifungal drugs is costly and time-consuming. As we’ll see, commercial interest in new antifungals has plateaued in recent years, which may exacerbate an already concerning situation as the threat of fungal diseases continues to evolve. New approaches are therefore needed, including advanced drug delivery systems, drug repurposing, and more.
Major classes of antifungals
The development of modern antifungals began in the mid-20th century with the introduction of polyenes. By the 1980s, azoles emerged as a safer and more available alternative to polyenes. The development of second-generation azoles, such as fluconazole (1990) and itraconazole (1992), marked a significant advancement. Fluconazole became widely used for treating candidiasis and cryptococcal meningitis, especially in HIV/AIDS patients.
The 21st century brought the introduction of echinocandins, a novel class of antifungal drugs that target fungal cell wall synthesis. These drugs offered a new mechanism of action and were effective against azole-resistant strains. The rise of antifungal resistance, particularly in Candida auris, has underscored the urgency of developing new treatments to counteract these constantly evolving threats. The FDA recently approved ibrexafungerp, the first drug in a newer class of antifungals called triterpenoids, for treating vulvovaginal candidiasis.
This approval marked a significant milestone in antifungal drug development, yet the emergence of antifungal resistance combined with the cost and time required for clinical trials are roadblocks to the development of new drugs. Below, we summarize the five major classes of antifungals on the market today — polyenes, azoles, echinocandins, pyrimidine analogs, and allylamines — including new developments:
- Polyenes are natural products consisting of large cyclic macrolides derived from Streptomyces bacteria. They bind to ergosterol in fungal cell membranes and act via a pore-forming model, resulting in compromised integrity of cell membranes. They are a broad-spectrum antifungal with a low propensity of resistance-development mechanisms, but they cannot be used for systemic fungal infections, except amphotericin B. They have limited water solubility leading to very low bioavailability. Recently, the new polyene derivative, mandimycin, which functions as an antifungal via targeting phospholipids instead of ergosterol, has demonstrated broad-spectrum fungicidal activity against many multidrug-resistant pathogens.
- Azoles are small molecules showing the presence of either a triazole or imidazole ring system. They disrupt ergosterol biosynthesis by inhibiting fungal cytochrome P450-dependent enzyme 14α-lanosterol demethylase (CYP51). Azoles’ improved bioavailability allows oral administration and treatment of systemic fungal infections, but resistance to these drugs has developed, resulting in decreased effectiveness. Recent developments with azoles include opelconazole completing its Phase II clinical trial in 2024; the ongoing Phase III clinical trial of opelconazole in combination with other systemic antifungals; and the FDA approval of oteseconazole in 2022.
- Echinocandins are semi-synthetic derivatives of natural products isolated from fungal strains. They function by inhibiting β-1,3-D-glucan synthase to compromise the integrity of the fungal cell wall. A broad-spectrum antifungal, echinocandins have minimal toxicity or instances of drug resistance, but they must be administered intravenously. In 2023, the drug Rezafungin was approved by the FDA.
- Pyrimidine analogs are antimetabolites that interfere with nucleic acid synthesis. Their target mechanism is to convert 5-Fluorocytosine to 5-fluorouridine monophosphate catalyzed by fungal enzymes, which leads to faulty protein synthesis when incorporated into fungal RNA. These drugs have a limited spectrum of activity and resistance to their antifungal properties is growing. However, new pyrimidine derivatives were identified and developed in 2024 using high throughput screening.
- Allylamines are a tertiary allylamine group that causes antifungal activity. They function by enzyme inhibition resulting in the accumulation of squalene and disrupted ergosterol synthesis, affecting the integrity of the fungal cell membrane. These drugs have broad-spectrum antifungal activity but resistance to them is reducing their effectiveness.
Mechanisms of antifungal resistance
The primary challenge in antifungal treatment is the emergence of drug resistance. This resistance is an evolving trait that allows fungi to survive despite the presence of damaging chemical agents. Across a period of profound global environmental change, overlap between plant and animal antifungal agents, and expanding at-risk populations, pathogenic fungi are evolving resistance to all licensed systemic antifungal drugs.
For example, the use of triazole fungicides in the U.S. increased four-fold from 2006 to 2016, and trends in azole fungicide use correlated with the sharp increase of azole-resistant Aspergillus fumigatus infections in humans. Fungicide use is expected to grow as a result of climate change and the ensuing need for more concentrated and frequent applications to compensate for productivity loss due to extreme weather.
The presence of indwelling catheters, artificial heart valves, and other surgical devices may also contribute to refractory infections, as infecting organisms attach to these objects and establish biofilms that resist drug action. Biofilms display an organized three-dimensional structure comprised of a dense network of yeast and filamentous cells embedded in an exopolymeric matrix of carbohydrates, proteins, and nucleic acids. Fungal biofilms exhibit higher tolerance to drugs due to several factors, including their extracellular polymeric substances matrix, slower growth rates, presence of persister cells, exchange of genetic material, and synergistic interactions in biofilm cells.
The most common forms of acquired resistance, however, involve mutations to target protein-binding sites. This mechanism is typical among azole-resistant strains of Candida species such as Candida albicans, Candida tropicalis, and Candida parapsilosis, as well as in azole-resistant Aspergillus species. Figure 2 summarizes the mechanisms of resistance in the five major classes of antifungals:

Research trends in antifungals reveal the need to identify new treatments
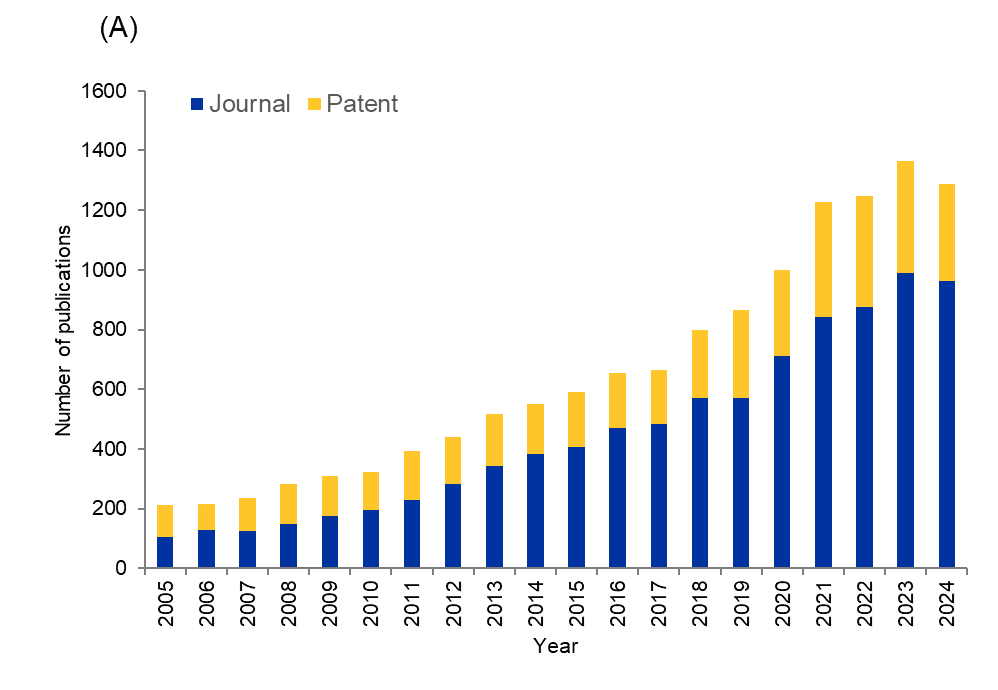
We analyzed the CAS Content CollectionTM, the largest human-curated repository of scientific information, to better understand how the scientific community is tackling the problem of fungal infections. We found that, while publications have grown over the last 20 years, numbers are plateauing and even declining — a concerning trend given the growth in resistance to existing antifungal medications (see Figure 3).
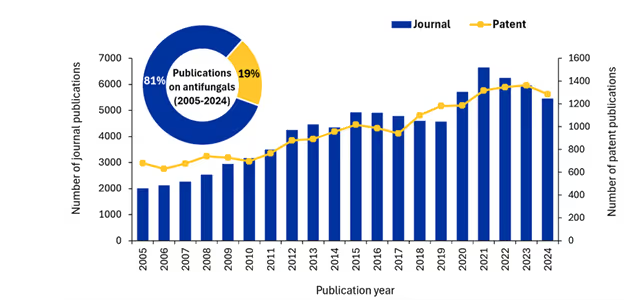
Journal publications show a steady upward trajectory from approximately 2,000 publications in 2005 to nearly 6,600 at their peak in 2021. We saw an initial gradual increase (2005-2012), a period of stabilization around 4,500-5,000 publications annually (2013-2019), and a dramatic surge reaching unprecedented levels in 2020-2021, followed by a moderate decline through 2024.
Patent publications follow a somewhat different trajectory while maintaining a general upward trend. The most recent years, however, show a slight decline in patents, which suggests that while academic interest has grown, there remain significant challenges to translating those discoveries into commercially viable antifungal therapies. The disparity is further emphasized by the distribution of research output, with patents constituting just 19% of total publications compared to journals' dominant 81% share.
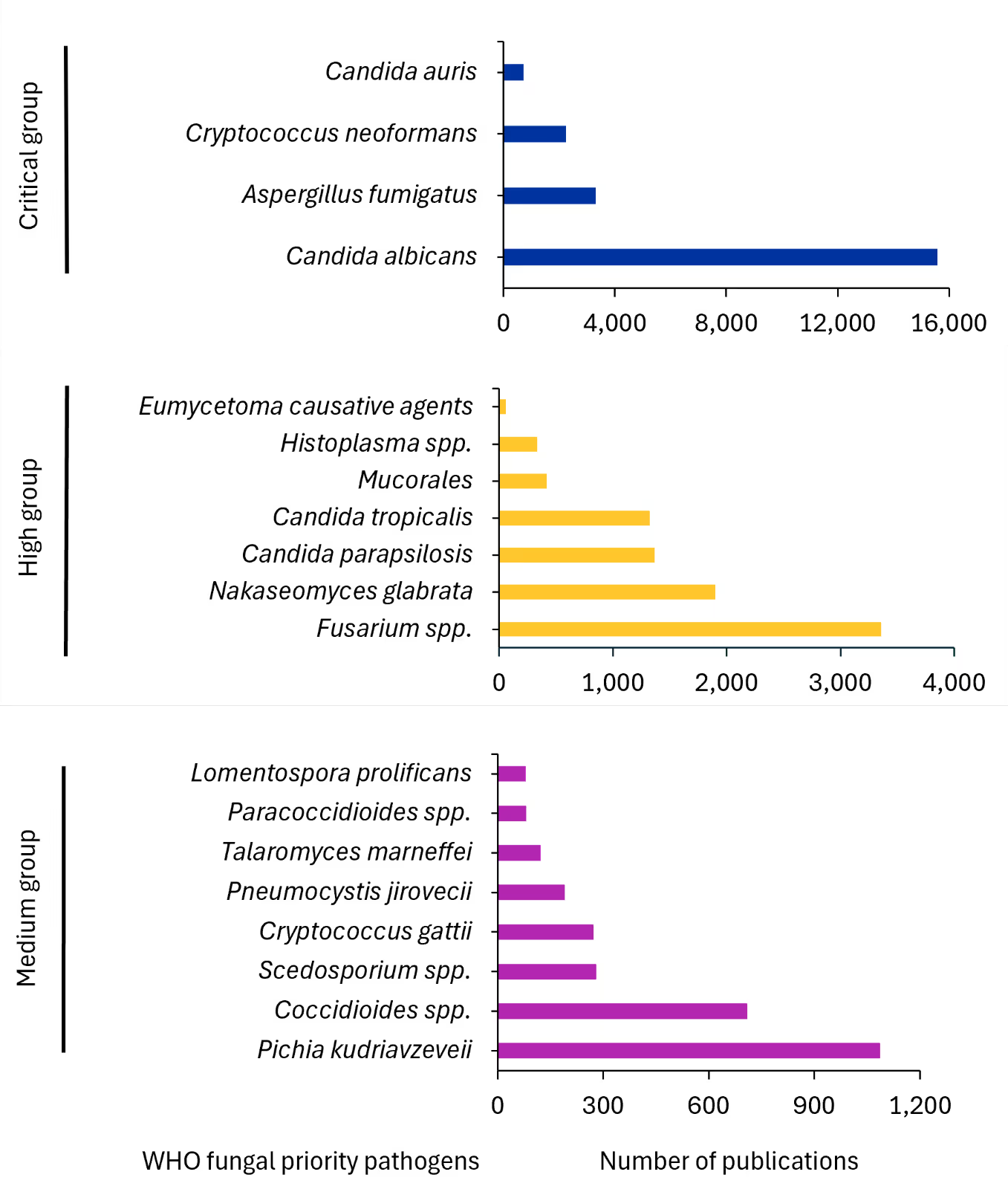
As noted, the WHO categorized priority fungal pathogens into three groups based on their public health impact and the need for research and development: critical, high, and medium priority. The critical priority fungal pathogens are the most urgent threats due to their high morbidity, mortality, and antifungal resistance. They primarily affect immunocompromised individuals, such as those with HIV/AIDS, cancer, or after organ transplants. They include Candida auris, Cryptococcus neoformans, Aspergillus fumigatus, and Candida albicans.
The high priority fungal pathogens also pose significant threats but are considered less urgent than the critical group. They cause severe infections and have emerging resistance patterns. They include Eumycetoma causative agents as well as Histoplasma, Fusarium, and several Candida species.
The medium priority fungal pathogens are important but currently have a lower public health impact compared to the critical and high-priority groups. They still require attention due to their potential to cause severe disease.
We analyzed the research attention given to various WHO-designated fungal priority pathogens across journal and patent literature from 2005-2024 (see Figure 4). Within the critical group, Candida albicans dominates research focus with approximately 16,000 publications, nearly four times the attention given to Aspergillus fumigatus (~4,000 documents), while Cryptococcus neoformans and Candida auris receive substantially less coverage despite their critical designation.
This analysis revealed that research is unevenly distributed across fungal threats, with certain pathogens like Candida albicans receiving attention while other WHO critical priority pathogens remain significantly understudied despite their public health importance.
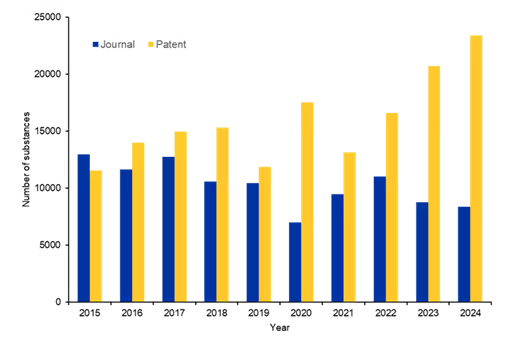
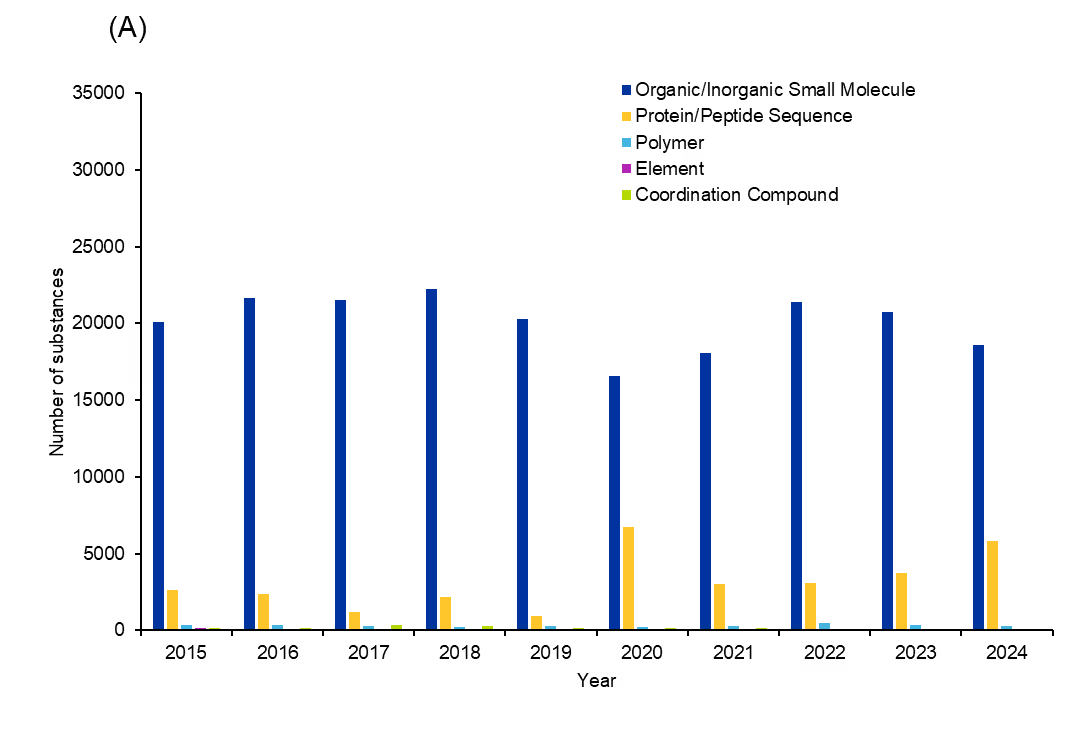
We further analyzed the last 10 years of publications from the CAS Content Collection to understand which substances are being explored in antifungals. Substance analysis was limited to relevant roles, including therapeutic (THU), pharmacological (PAC), or pharmacokinetic (PAK). The number of substances per year shows an increasing trend from the year 2021 onwards (see Figure 5).
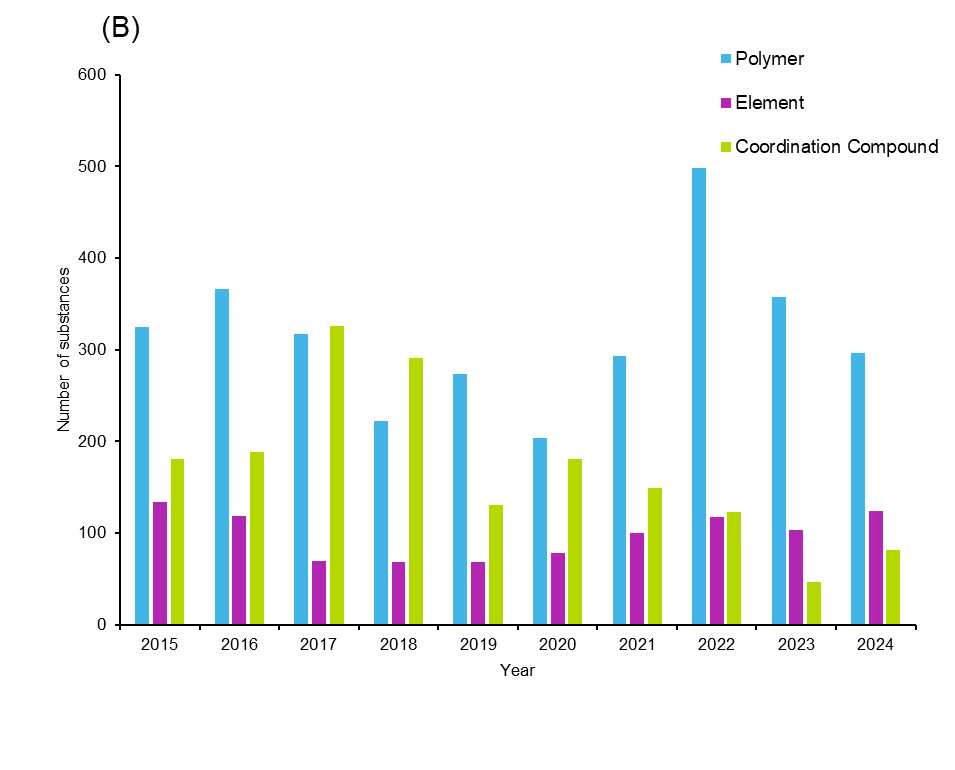
Further investigation into the substance classes indicates that organic and inorganic small molecules, protein/peptide sequences, polymers, elements, and coordination compounds are the major classes explored in antifungals (see Figure 6).
Polymers are an interesting category of substances because polymers or polymer-based hydrogels can be used as drug delivery agents or they themselves can act as antifungals. Natural polymers like chitosan and cellulose, and synthetic polymers like polyvinyl alcohol, polyethylene glycol, and poly acrylic acid, are most common in antifungals. While these substances account for a small fraction of substances, we found a trend of increasing interest that peaked in 2022. This suggests that unconventional substances and materials are being eyed more intentionally for antifungal therapy.
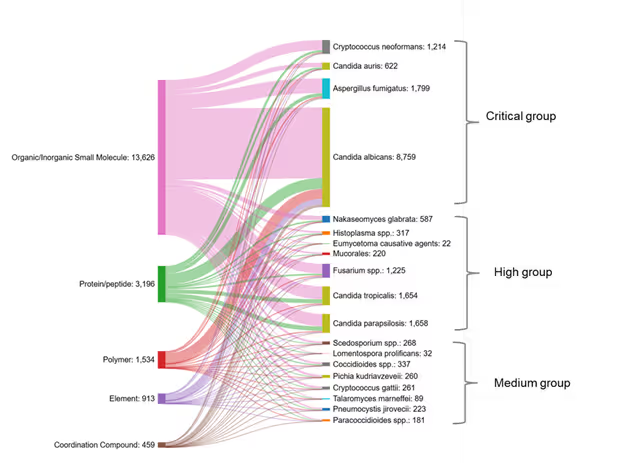
We also examined the co-occurrences between various substance classes and different fungal species in the literature (see Figure 7). The fungal species are arranged here according to the WHO classification: critical group, high group, and medium group. Candida albicans has the maximum number of substances co-occurring with it, followed by Candida tropicalis, Candida parapsilosis, and Aspergillus fumigatus.
Organic/inorganic small molecules co-occur most often with Candida species and Aspergillus fumigatus. Three pathogens from the high-priority group and two pathogens from the medium-priority group have very few substances co-occurring with them (less than 100), indicating the need for identifying novel agents which can act against these pathogens.
Clinical trials slowing down
If publications related to fungal pathogens are plateauing or even declining, what does that mean for preclinical and clinical trials for antifungal treatments? When examining preclinical candidates’ activity against the WHO priority groups of fungal pathogens, it is obvious that most drugs have activity against multiple fungal species (see Table 1). For example, in the critical group, 19 drugs have activity against Candida albicans, 15 against Candida auris, 12 against Aspergillus fumigatus, and eight against Cryptococcus neoformans.
Table 1: WHO priority group levels, fungal pathogens, and the number of preclinical candidates with activity against them. Source: CAS, publicly available information.
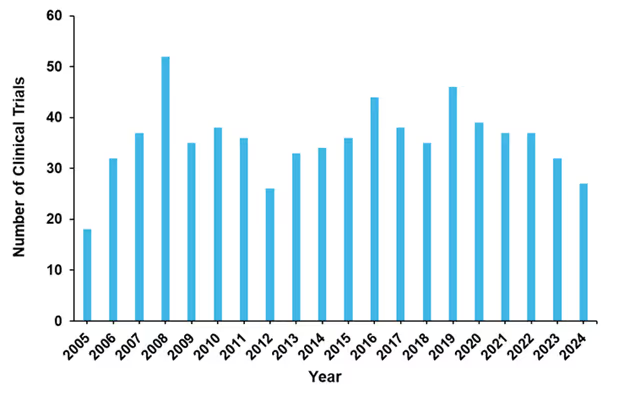
In terms of clinical trials, the majority (71%) taking place in the last 20 years are completed. In Figure 8, we can see that the number of trials has oscillated over the years, with a current downward trend. Over the past decade, only eight antifungal drugs have been approved by the U.S. FDA. Only four drug candidates are currently in Phase III clinical trials.
Of the nearly 100 active clinical trials in the pipeline, only 18% are evaluating novel agents rather than a repurposed or reformulated drug. As seen in Table 2, 12 of these drugs are small molecules, five are biologic peptide therapies, and one is a live microorganism.
|
Intervention |
Sponsor, location |
Active against |
|||
|
A. fumigatus |
C. auris |
C. albicans |
C.neoformans |
||
|
Azoles |
|||||
|
†HCP-02 (Phase I) |
HC Synthetic Pharmaceutical, China |
y |
y |
y |
n |
|
†VT 1598 (Phase I/NCT04208321) |
Mycovia Pharmaceutical, United States |
y |
y |
y |
y |
|
†Opelconazole (Phase III/NCT05238116) |
Pulmocide, United Kingdom |
y |
n |
y |
n |
|
Peptide-based |
|||||
|
BAL-2062 (Phase I) |
Basilea Pharmaceuticals, Switzerland |
y |
n |
n |
n |
|
Occidiofungin B (Phase I) |
Sano Chemicals, United States |
n |
y |
y |
n |
|
§PL-18 (Phase I/NCT05340790) |
Protelight Pharmaceutical, China |
n |
y |
y |
n |
|
Novexatin (Phase II) |
NovaBiotics, United Kingdom |
n |
y |
y |
n |
|
**Pezadeftide (Phase II) |
Hexima, Australia |
n |
y |
y |
n |
|
Polyenes |
|||||
|
§SF-001 (Phase I) |
Elion Therapeutics, United States |
y |
n |
n |
n |
|
*BSG005 |
Biosergen, Sweden |
y |
y |
y |
y |
|
Echinocandin |
|||||
|
*HRS9432 (Phase II/NCT06194201) |
Jiangsu Hengrui Pharmaceuticals, China |
n |
y |
y |
n |
|
Gepix |
|||||
|
§Fosmanogepix (Phase III/NCT05421858) |
Amplyx Pharmaceuticals, United States |
y |
y |
y |
n |
|
Orotomide |
|||||
|
†Olorofim (Phase III/NCT05101187) |
F2G, United Kingdom |
y |
n |
n |
n |
|
Triterpenoid |
|||||
|
†SCY-247 (Phase I) |
Scynexis, United States |
y |
y |
y |
n |
|
Others |
|||||
|
Zambon antifungal (Phase I) |
Zambon Pharmaceutical, Italy |
Activity not reported |
|||
|
§ATB1651 (Phase II/NCT06327295) |
AmtixBio, Korea |
n |
n |
n |
n |
|
BGY-1601 (Phase II/NCT06450990) |
Nexbiome Therapeutics, France |
n |
y |
y |
n |
|
WXSH0102 (Phase II/NCT06771063) |
Cisen Pharmaceutical, India |
n |
y |
y |
n |
|
TY-1801J (Phase III/CTR20231279) |
Kaken Pharmaceutical, China |
Activity not reported |
|||
†fungal cell wall synthesis inhibitor
§fungal cell wall membrane disruptor
*fungal cell wall disruptor
**mitochondria disruptor
Table 2: Active antifungal agents in the clinical trial pipeline with their clinical trial phase, class/mode of action, CAS RN, sponsor, identifier, and WHO critical fungal activity. Source: CAS, publicly available information.
Researchers exploring more treatment approaches for new antifungals
The rise in drug-resistant fungi combined with environmental changes and infection opportunities via healthcare settings is problematic. The fact that these factors are converging at a time when research publications and clinical trials for antifungal treatments are slowing down is equally troubling. However, this also means there are significant opportunities for new breakthroughs.
As our analysis shows, there are many promising options for new therapeutics, ranging from re-examining existing drugs for their antifungal efficacy to cutting-edge gene therapy and drug delivery systems. Figure 9 summarizes these approaches:
Identifying newer fungal targets
Traditional antifungal therapies often target the fungal cell wall or membrane, such as azoles targeting ergosterol synthesis or echinocandins inhibiting β-glucan synthase. A critical area of new research is identifying novel targets in fungal pathogens to develop drugs against those targets.
- Targeting the fungal cell wall polysaccharides:
- β (1,6)-glucan synthesis: β(1,6)-glucan is a crucial component of fungal cell walls. However, our limited knowledge of β (1,6)-glucan synthesis has complicated the development of antifungals targeting this component. The pyridobenzimidazole derivative D75-4590 (CAS RN 384376-42-5) was shown to block the function of Kre6 in Candida albicans (one of the important genes in β(1,6)-glucan synthesis).
- Chitin biosynthesis: There is covalent bonding between chitin and β (1,3)-glucan to form the cell wall structural layer, making it an appealing drug target. However, structural and functional redundancies for chitin synthase (Chs) have complicated the development of chitin biosynthetic targets. Nikkomycins are natural peptides that inhibit Chs enzymes and have shown antifungal activity. Other compounds, such as novel 3-substituted amino-4-hydroxycoumarin derivatives, exhibited moderate to excellent inhibitory activity against Chs protein.
- Targeting the fungal cell wall proteins:
- Glycosylphosphatidylinositol (GPI) anchor synthesis: The GPI anchor is a glycolipid moiety that links many cell wall proteins to the plasma membrane. The biosynthesis of GPI anchors involves a complex pathway with multiple enzymatic steps. Gwt1, a mannosyltransferase involved in the early stages of GPI anchor biosynthesis, is the target of fosmanogepix (E1210/APX001, CAS RN 2091769-17-2), a novel broad-spectrum antifungal agent that has demonstrated efficacy against Aspergillus and other filamentous fungi and is in late clinical phase of development.
- Mannoprotein biosynthesis: Mannoproteins, heavily glycosylated proteins, form the outermost layer of the yeast cell wall and play crucial roles in adhesion, biofilm formation, and interaction with the host immune system. Inhibiting their synthesis or modification could disrupt cell wall integrity and expose underlying cell wall components to antifungal drugs or the host immune system. Pradimicins bind mannan and disrupt the cell wall of fungi, and they exhibit low toxicity in vivo. A pradimicin derivative, BMS-181184 (CAS RN 139272-69-8), has shown in vitro activity against Candida neoformans, Aspergillus species, other Candida species as well as molds.
- Targeting the fungal cell membrane:
- Ergosterol biosynthesis: Azole antifungals inhibit the cytochrome P450 enzyme CYP51, leading to the depletion of ergosterol and the accumulation of toxic sterol intermediates, disrupting membrane function. Novel azoles, such as VT 1598 (CAS RN 2089320-99-8), are being developed to overcome resistance to known azoles. This compound also exhibits improved selectivity for fungal CYP51 over human cytochrome P450 enzymes, potentially reducing drug-drug interactions and toxicities.
- Glycosphingolipid synthesis: Glycosphingolipids are important components of fungal membranes involved in cell signaling, growth, and virulence. Inhibiting enzymes like inositol phosphorylceramide synthase, which is absent in humans, could provide a selective antifungal strategy.
- Proton ATPases: These membrane-bound enzymes maintain the electrochemical gradient across the plasma membrane, essential for nutrient uptake and cell survival. Fungal-specific features of these ATPases are being explored for selective inhibition.
- Targeting intracellular processes: Several intracellular pathways and processes that are essential for fungal survival and virulence differ significantly from their mammalian counterparts, representing promising targets for antifungal drug development:
- Mitochondrial respiration
- Siderophore uptake
- Pyrimidine biosynthesis
- Riboflavin biosynthesis
- Glyoxylate cycle
- Calcineurin pathway
- Protein folding
- Amino-acid synthesis pathway
Improving existing drugs
- Novel or better formulations: Creating intravenous (IV) drug formulations already available in oral formulation is a key strategy. IV formulations have better bioavailability and are useful in cases where the infection is severe. For example, Posaconazole, which was initially available as an oral suspension, has been reformulated into tablet and IV forms.
- Combination therapy: Another approach in fighting the rising antifungal resistance and the limited arsenal of antifungal drugs is the use of synergistic combinations of existing antifungals. Synergistic combinations occur when two or more drugs work together to produce an effect greater than the sum of their individual effects. This strategy not only enhances antifungal efficacy but also reduces the likelihood of resistance development. Combining drugs that act on different fungal pathways can disrupt multiple essential processes simultaneously, making it harder for fungi to develop resistance. One such example includes azoles combined with echinocandins.
Drug repurposing
With the rise of multidrug-resistant fungal pathogens and the limited pipeline of new antifungal drugs, repurposing existing drugs for antifungal activity offers a promising alternative. Since repurposed drugs have already undergone extensive safety testing, the time and cost required for preclinical and early clinical development are significantly reduced. Also, the pharmacokinetics, toxicity, and side effects of existing drugs are well-documented, minimizing the risk of unexpected adverse effects. Some examples of repurposed drugs with antifungal activity include antidepressants like Sertraline, anticancer drugs like Tamoxifen, and antibiotics such as Doxycycline.
For many of the drugs which are being repurposed, the precise fungal targets are unknown, hence further studies are required to identify those. Techniques like transcriptomic analysis and advanced genetic assays will help discover the targets and establish the foundation for developing novel antifungal drugs.
Artificial intelligence
Artificial Intelligence (AI) has emerged as a powerful tool in antifungal research, offering the potential to accelerate the identification of novel antifungal compounds, optimize drug design, and predict resistance mechanisms. By leveraging machine learning, deep learning, and other AI techniques, researchers can analyze vast datasets, uncover hidden patterns, and make predictions that would be impossible using traditional methods.
- Predictive modeling: AI-based tools can help identify new antifungal compounds and understand drug-target interactions. Some examples include using artificial neural networks for predicting the antifungal properties of quaternary ammonium salts against Candida albicans. Additionally, predictive tools such as AlphaFold 3 (AF3) are expected to enrich the structural biology of antifungal targets.
- High-throughput screening: Machine learning, especially deep learning, has demonstrated significant advantages in drug discovery and development by accelerating high-throughput screening. A recent study identified drugs that can be repurposed as antifungals with high confidence using machine learning-based methods.
- De-novo drug design: Generative models, including deep generative adversarial networks (GANs), have shown promising results in designing de novo drugs by creating novel molecular structures with desired properties. AI has helped significantly in speeding up the development of antifungal peptides (AFPs) by combining deep learning approaches with QSAR studies.
- Analysis of omics data: AI can analyze fungal genomic, transcriptomic, and proteomic data to identify potential drug targets and resistance mechanisms. Omics data is usually complex and bulky, posing several challenges that can be addressed by AI. Some applications of AI in analyzing omics data include reducing data variability, noise reduction, and data integration.
CRISPR/Cas-based gene editing
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated (Cas) proteins have transformed genetic engineering, offering unprecedented precision, efficiency, and versatility. Originally discovered as part of the bacterial immune system, CRISPR-Cas technology has been adapted for use in many organisms, including fungi. The most used CRISPR-Cas9 system for genome editing in fungi is the type II Cas9 system, which is recognized for its high specificity and efficiency.
There are, however, several limitations to CRISPR/Cas technology for fungal gene editing. Fungi have complex cell walls and structures that can make the delivery of gene-editing tools challenging. Different fungal species require tailored approaches for codon optimization, promoter selection, and delivery systems. For example, in many Candida species, the nucleotide sequence CTG is for human cells are not efficient in these species.
Fungal species also show variation in their DNA repair mechanisms, relying on either homology-directed repair (HDR) or non-homologous end joining (NHEJ). This variability affects the efficiency of gene editing tools. The stability and integration of plasmids and vectors can be challenging. Some species require linearized vectors (i.e., Candida albicans and Crytococcus neoformans), while others benefit from episomal plasmids or ribonucleoprotein complexes (Candida glabrata, Aspergillus fumigatus, and Mucorales).
CRISPR/Cas based gene editing in fungi is still at preliminary stages and currently there are no clinical studies evaluating their potential in the treatment of fungal infections. However, multiple preclinical studies have expanded our understanding of fungal pathogenicity and resistance mechanisms. Morio et al. showed that in Candida orthopsilosis, certain CoERG11 mutations confer resistance to azoles. Rybak et al. demonstrated that overexpression of CDR1 is a significant contributor to clinical triazole resistance in Candida auris. In another study, genes contributing to fungal survival under the stress of echinocandins were identified using CRISPR/Cas based gene editing in Aspergillus fumigatus.
Future research on the application of CRISPR/Cas in fungi will continue to provide useful insights leading to a more comprehensive understanding of this complex genus and eventually help in developing novel antifungals.
Antifungal peptides
Antifungal peptides (AFPs) are small cationic peptides that exhibit strong antifungal activities against several pathogenic fungi, making them potential candidates for developing novel antifungal agents. AFPs exhibit several advantages over conventional antifungals, like rapid killing kinetics, lower tendency for resistance development, and diverse mechanisms of action. AFPs are most commonly classified based on their source — natural, semi-synthetic, and synthetic. APD3 is a widely used database used for research purposes which enlists antimicrobial peptides originating from various sources. As of April 2025, there were 1,550 antifungal peptides reported, with 932 anti-candida peptides.
- Natural peptides: Natural AFPs were discovered by testing their antagonistic activity in vitro against pathogenic fungi. However, newer methods such as template-based, docking simulations, and other sequence-based methods are becoming more common for novel in silico prediction of AFPs. We analyzed the publications present in CAS Content Collection related to antifungal therapeutics and identified defensins as the most-studied natural AFP.
- Semi-synthetic and synthetic peptides: The aim of creating semi-synthetic and synthetic peptides is to improve pharmacological properties like stability and bioavailability, reducing toxicity and/or the immunogenicity of natural peptides. Structure-activity relationships (SAR) are crucial for the design and development of these AFPs.
Immunotherapy
The complex interaction between the host immune system and invading fungal pathogens dictates the outcome of infection. This understanding has encouraged the exploration of immunotherapy approaches for treating fungal infections. Various immunotherapeutic modalities like cytokine therapies, checkpoint inhibitors, adoptive cell transfer, and vaccines are being explored as potential antifungal treatment options. The aim of using these is to modulate host immune response to enhance clearance of fungal pathogens and thereby improving treatment outcomes.
Newer drug candidates
There are numerous antifungal classes outside of the main five that are being researched, some of which were discovered decades ago, and some that are new. For example, myriocin was first discovered in the 1970s, but is being studied in combination with known antifungals such as flucanazole. Researchers are exploring natural products like Khafrefungin and Rustmicin, and small molecules like PQA-Az-13 have also shown promising results.
Innovative drug delivery systems can improve antifungal efficacy
Delivery vectors for antifungal drugs are systems or methods used to transport antifungal agents to the site of infection in the body. These vectors aim to improve efficacy, reduce side effects, and enhance the targeted delivery of antifungal drugs. The specificity of delivery vectors for antifungal drugs is crucial for maximizing therapeutic efficacy while minimizing side effects. Different drug delivery systems (DDS) are tailored to the physicochemical properties of the antifungal drugs and the nature of the fungal infection.
- Liposomes: These are spherical vesicles comprising a phospholipid bilayer that can encapsulate hydrophilic and hydrophobic drugs. Their advantages include enhanced drug solubility and stability, reduced toxicity by targeting specific tissues, and prolonged drug release. Exemplary antifungal drugs in liposomal formulations include amphotericin B, fluconazole, and nystatin.
- Nanoparticles: These are tiny particles (1-100 nm) that can be made from various materials, including lipids, polymers, and metals. Their advantages are improved drug delivery to specific sites, enhanced bioavailability, controlled release, and reduced side effects. Exemplary fungicides delivered by nanoparticles include fluconazole, itraconazole, and voriconazole. Solid lipid nanoparticles are a promising class of submicron-sized particles that show improved stability as well as reduced toxicity and enhanced targeting. Carbon nanotubes (CNTs) are another promising platform for antifungal drug delivery.
- Micelles: These are self-assembling structures formed by amphiphilic molecules in aqueous solutions. Their main advantages include increased solubility of hydrophobic drugs, targeted delivery, and reduced systemic toxicity.
- Niosomes: These are non-ionic surfactant-based vesicles similar to liposomes. Their advantages are enhanced stability compared to liposomes as well as controlled and sustained drug release.
- Dendrimers: These are highly branched, tree-like structures with a high degree of surface functionality. They exhibit high drug-loading capacity, precise targeting, and controlled release.
- Microemulsions: These are stable, isotropic mixtures of oil, water, and surfactant. They exhibit enhanced drug solubility and absorption and improved bioavailability.
- Hydrogels: These are three-dimensional networks of hydrophilic polymers that can swell in water. They are suitable for topical and mucosal applications, for sustained and localized drug release.
- Cyclodextrins: These are cyclic oligosaccharides that can form inclusion complexes with drugs. They enhance solubility and stability of drugs, reduce irritation, and improve bioavailability.
Antifungal materials address healthcare-associated infections
Fungal infections in healthcare settings, often termed healthcare-associated infections, are a serious concern. As noted, invasive medical procedures and the use of implanted medical devices create entry points and surfaces for fungal pathogens to colonize and form biofilms. Candida albicans, Aspergillus fumigatus, and other opportunistic fungi are common culprits, particularly in immunocompromised individuals.
The development of antifungal biomaterials aims to combat these infections by preventing initial fungal adhesion and biofilm formation or by delivering antifungal agents directly at the site of potential infection. These materials represent a growing field at the intersection of materials science and biomedical engineering, offering innovative solutions to combat fungal infections.
We analyzed the CAS Content Collection related to antifungal materials for the past 20 years. There is a consistent increase in journal and patent publications (see Figure 10). However, the growth in documents is closer to 10-fold as compared to that for patents, which is only around three-fold. We further identified the top fungal pathogens mentioned in these publications. As shown in Figure 10B, Candida albicans was the most common pathogen, followed by three Aspergillus species, and lastly Candida glabrata.

Conventional antifungal materials which have been used widely include nanoparticles, polymers, and hydrogels. Among nanoparticles, metals are among the most extensively studied, such as silver nanoparticles that disrupt fungal cell membranes, interfere with metabolic processes, and generate reactive oxygen species.
Polymers, both natural and synthetic, form the backbone of many biomaterials. Incorporating antifungal agents or modifying the polymer structure to impart antifungal properties is a common strategy. Polymers can be loaded with antifungal drugs and used to treat chronic superficial infections. Hydrogels are another emerging form of biomaterials. Their biocompatibility, flexibility, and ability to encapsulate drugs make them ideal for wound dressings and catheter coatings.
The field of antifungal biomaterials is constantly evolving with the exploration of novel materials with unique properties, for example metal-organic frameworks (MOFs), quantum dots, ionic liquids, and bioactive glasses. These innovations could signal important treatment breakthroughs, but long-term efficacy and safety still need to be evaluated in clinical trial settings. The fabrication of these materials must also be cost-effective and scalable to bring these innovations into wider clinical usage.
Future directions for antifungal treatments
Antifungal drug development is rapidly changing due to the increasing prevalence of fungal infections, the rise of drug-resistant strains, and the limited arsenal of effective antifungal agents. Despite exploring newer approaches such as identifying fungal-specific targets, immunotherapy, combination therapy, and more, the clinical development of antifungal therapeutics has been slow. Over the past decade, only eight antifungal drugs have been approved by the FDA, and only four drug candidates are currently in Phase III clinical trials.
This is attributed to several challenges, notably high development costs, complex host-fungal interactions, and the need to understand emerging pathogens in a warming climate. The future of antifungal therapy will therefore be a multifaceted approach that combines novel drug discovery, advanced technologies, and a deeper understanding of fungal biology and host immunity.
Innovative strategies, such as immunotherapy, nanotechnology, and AI-driven drug discovery, hold great promises for addressing the growing burden of fungal infections. Collaborative efforts between researchers, clinicians, and pharmaceutical companies are essential to overcome current challenges and improve patient outcomes.
To learn more about antifungal therapeutics, see our publication at a pre-print server.




